

С.А. ЗЯТЬКОВ, Г.Г. ГОНЧАРЕНКО
Разработаны и апробированы методы молекулярно-генетической идентификации Felis catus акромеланистических окрасов, позволяющие четко выявлять гетерозиготных особей (генотип Ccs), которые фенотипически неотличимы от особей с генотипом СС.
Ключевые слова: домашняя кошка, Felis catus L., пигментогенез, аллель cs, тирозиназа.
The methods of molecular genetic identification of Felis catus of acromelanistic colors have been developed and tested, which makes it possible to clearly identify heterozygous individuals (Ccs genotype) that are phenotypically indistinguishable from individuals with the CC genotype.
Keywords: domestic cat, Felis catus L., pigmentogenesis, allele cs, tyrosinase
Несмотря на многовековую историю взаимоотношений домашней кошки (Felis catus L.)
с человеком, она практически не изменилась и по внешнему виду похожа на предков (F.
libyca Forster, F. chaus Güldenstaedt, F. sylvestris Schreber). Следует согласиться с мнением
многих исследователей, что современную домашнюю кошку, за исключением некоторых декоративных пород, нельзя считать в полном смысле домашней, поскольку она сохраняет в
неизменном виде привычки и поведение своих диких сородичей, лишь приспособив их к условиям жизни вблизи с человеком. С другой стороны, долгое сожительство F. catus рядом с
человеком способствовало смягчению действия естественного отбора и тем самым благоприятствовало возникновению и закреплению большого числа легко идентифицируемых по
внешнему виду животных мутантных генов окраски, чего никогда не наблюдают в популяциях диких животных. Все это обеспечило накопление такого множества расцветок меха у
F. catus, которое мы сейчас можем наблюдать [1], [2].
Необходимо отметить, что формирование окраса меха у кошек начинается в процессе
пигментогенеза (меланогенового пути) [3]–[6], который является генетически детерминированным и начинается еще на эмбриональной стадии. На очень ранних этапах развития у зародыша в области нервной трубки выделяется закладка будущих пигментных клеток меланоцитов. Сами они производить пигмент еще не могут, для этого им нужно претерпеть ряд
изменений. Прежде всего, эти клетки, так называемые меланобласты, должны принять веретенообразную форму, пригодную для миграции. Мигрируют они сначала в центры пигментации, а оттуда – в волосяные фолликулы [1], [3]. Процесс формирования меланоцитов из
клеток нервного валика представлен на рисунке 1.
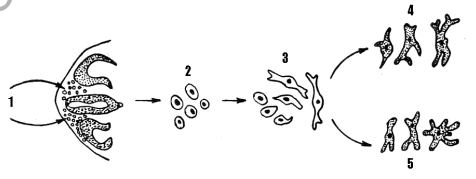
Рисунок 1 – Миграция меланобластов из нервного валика [3]:
1 – клетка нервного валика, 2 – меланобласты, 3 – незрелые меланоциты, 4 – эпидермальные меланоциты, 5 – дермальные меланоциты
Гранулы пигментов, носящие название меланосом, образуются в меланоцитах. У большинства видов млекопитающих меланоциты имеют типичное строение, связанное с наличием в клетке отростков и равномерным распределением гранул меланина.
Окраска меха у кошек определяется присутствием пигмента – меланина, который в зависимости от химического состояния может находиться в двух формах эумеланина и феомеланина. Гранулы эумеланина сферические и поглощают почти весь свет, давая черную пигментацию, в то время как гранулы феомеланина продолговатые (эллипсоидной формы) и отражают свет в красно-желто-оранжевом диапазоне [1], [3]–[7]. Этапы биосинтеза пигментов у кошек из аминокислоты тирозина и некоторые ферменты, участвующие в метаболизме, представлены на рисунке 2.

Пути синтеза эумеланина и феомеланина из тирозина
(отмечена множественная роль тирозиназы)
Та или иная окраска животного является конечным результатом сложного процесса
формирования и функционирования структур, связанных с образованием пигмента. Каждый
из этих этапов пигментогенеза имеет соответствующий генетический контроль. При нормальном функционировании механизма биосинтеза пигментов и их распределении в волосяном покрове кошек формируется «окрас дикого типа» (серый полосатый). Все другие цветовые вариации в окрасе меха у кошек обусловлены мутациями в соответствующих генах, ответственных за формирование и распределение пигментов [1].
Целью работы было отработать методику молекулярно-генетической идентификации
особей F. catus акромеланистического окраса.
Акромеланистический окрас проявляется из-за неравномерного окрашивание тела кошки. Такие животные имеют темную голову, конечности и хвост и значительно более светлый
корпус. Эти окрасы возникают в результате наличия температуро-чувствительной формы тирозиназы. При нормальной температуре тела активность данной формы тирозиназы резко
снижена, что и приводит к осветлению окраса. Сниженная температура головы и конечностей способствует активации фермента и запускает нормальный синтез пигмента, что и
обеспечивает развитие типичного «сиамского» окраса (рисунок 3) [1], [8]. За формирование
данного признака отвечает рецессивный аллель сs локуса Color.
Локус Color имеется у всех млекопитающих и определяет работу регуляторного белкафермента – тирозиназы. Наличие в генотипе доминантного аллеля С этого локуса обеспечивает синтез нормальной тирозиназы, и тогда процесс наработки не только пигментов в меланоцитах, но и дофамина и ДОФА в других клетках нейрального происхождения протекает в
полном объеме (рисунок 2). У кошек в этом локусе помимо нормального доминантного аллеля С, который отвечает за нормальный синтез пигментов, имеется еще серия рецессивных
мутантных аллелей (C > cb= cs > c, cm) [1], [9]–[12].

Рисунок 3 – Сиамские кошки (генотип cscs)
Необходимо отметить, что долгое время оставалась неизвестной локализация аллелей
локуса Color в хромосоме, а также их нуклеотидная последовательность и молекулярногенетический механизм проявления акромеланистического окраса. Разрешение этих вопросов стало возможным после успешных исследований в рамках проекта «Геном кошки»
(2002–2007) [13]–[14]. Было установлено, что в локусе Color (C) у млекопитающих находится ген, кодирующий тирозиназу (TYR) – фермент, участвующий в синтезе меланинов. Оказалось, что для таких пород кошек, как бирманская (соболиный окрас, генотип cbcb), сиамская (окрас колор-поинт, генотип cscs) (OMIA 000202-9685) в пигментогенезе принимает участие температуро-чувствительная тирозиназа. Приведенные выше породы были обнаружены
в Таиланде и известны там как Суфалак (Suphalak) – бирманская и Вичиен-маат (Wichienmaat) – сиамская. Кроме того, в гене TYR обнаружены два полных варианта альбино (аллелиc, cm) (OMIA 000202-9685). Аллель с в гомозиготном состоянии (генотип сс) вызывает формирование сплошного белого окраса. Это явление происходит из-за полной инактивации фермента тирозиназы приводит к альбинизму. Кошки с данной мутацией фенотипически схожи с животными с генотипом W- (ген White). Различия между ними заключается в окраске зрачков, которые у кошек-альбиносов с генотипом сс – розовые или красные из-за полного отсутствия пигмента в радужной оболочке [1]. Кроме того, селекционная работа, проводимая с кошками бирманской породы, позволила выявить необычный окрас, названный «мокко» («mocha»), генотип cmcm. Кошки с окрасом «мокко» имеют аквамариновую окраску глаз, розовую кожу носа и подушечки лап [12].
Помимо этого, известно много примеров альбинизма причиной которых являются мутации по гену TYR. Например, варианты с потерей функции, которые вызывают кожноглазной альбинизм типа 1A (OCA1A; OMIM: 203100) или варианты, которые снижают ферментативную активность, вызывая кожно-глазной альбинизм типа 1B (OCA1B; OMIM:606952) у людей [12].
Таким образом, было показано, что ген тирозаназы F. catus находится в D1 хромосоме,
а кодирующая его часть имеет размер 1587 н.п. и включает 5 экзонов (GenBank: AY959314–
AY959318) (рисунок 4) [10], [15].
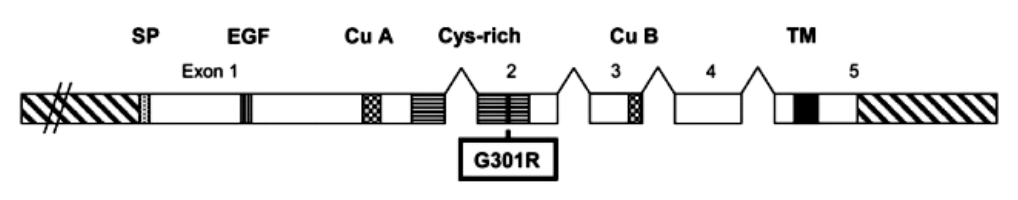
Рисунок 4 – Структура гена тирозиназы: SP – сигнальный пептид; EGF – домен фактора эпидермального роста; Cu A и Cu B – медьсвязывающие домены; Cys-rich – домен, богатый цистеином; TM – трансмембранный домен
На рисунке указано место локализации несинонимичной мутации во 2 экзоне (TYR901G>A), приводящий к замене глицина (G) на аргинин (R) в положении 301 кодона (G301R).
Даная мутация определяет у особей с генотипом cscs акромеланистичный окрас (рисунок 3).
В этой связи особи с сиамским и обычным «несиамским» окрасом отличаются друг от
друга однонуклеотидным замещением (SNP) во втором экзоне гена тирозиназы. Нуклеотидная последовательность фрагмента ДНК, содержащего 2 экзон гена тирозиназы для особей с
аллельным вариантом сиамского типа (аллель cs) (GenBank: AY743348.1) и особей «несиамского» дикого типа (аллель С) (GenBank: AY743346.1) представлены на рисунках 5 и 6 [9].

Рисунок 5 – Фрагмент ДНК, содержащий 2 экзон гена тирозиназы для особей с аллельным
вариантом сиамского типа (аллель cs)

Рисунок 6 – Фрагмент ДНК, содержащий 2 экзон гена тирозиназы для особей с аллельным
«несиамского» типа (аллель С)
На предложенных рисунках 5 и 6 хорошо заметно месторасположение однонуклеотидного замещения (выделено цветом), кроме того на этом же рисунке подчеркнуты участки ДНК
для пары праймеров, которые успешно используются при амплификации аллелей локуса TYR.
Таким образом, последовательность праймеров необходимая для амплификации предложенного фрагмента размером 179 н.п. будет следующей:
– прямой праймер – tagccgattggaggatacaa;
– обратный праймер – gcagctttatccatggaacc.
Фрагменты ДНК размером 179 н.п., полученные после амплификации, присутствуют как у особей F. catus «симаского», так и «несиамского» окраса. Поэтому для их идентификации была использована рестиктаза Hpa II (рисунок 7). Эта рестриктаза имеет следующий сайт узнавания (стрелками указаны сайты рестрикции):
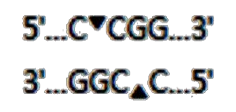
Рисунок 7 – Сайта узнавания и рестрикции для рестиктазы Hpa II
Сайт узнавания для предложенной рестиктазы как раз и содержит наше однонуклеотидное замещение. Это хорошо заметно на рисунках 8 и 9.

Рисунок 8 – Фрагмент ДНК, содержащий сайт узнавания для Hpa II у особей с аллельным
вариантом сиамского типа (аллель cs)

Рисунок 9 – Фрагмент ДНК, содержащий сайт узнавания для Hpa II у особей с аллельным
вариантом несиамского типа (аллель С)
В этой связи после рестрикции у особей «сиамского» окраса фрагмент размером 179 н.п. останется не тронутым, а у особей «несиамского» окраса будет подвержен рестрикции с образованием фрагментов размером 62 н.п. и 117 н.п.
Ампликоны, полученные в ходе ПЦР после рестрикции, в дальнейшем подвергали
гель-электрофорезу с флуоресцентным интеркалярным красителем бромистым этидием. Визуализация процесса электрофореза осуществлялась с помощью трансиллюминатора, который дает ультрафиолетовое излучение с длиной волны 365 нм, оптимальное для фиксации
результатов, полученных в результате ПЦР-анализа.
Таким образом, электрофорез в агарозном геле позволяет разделить, а затем легко извлечь полученные в результате ПЦР фрагменты ДНК в чистом виде для последующего использования. Кроме того, анализируя электрофоретические спектры ДНК на геле, можно точно установить генотип F. catus по гену, кодирующему тирозиназу (рисунок 10).
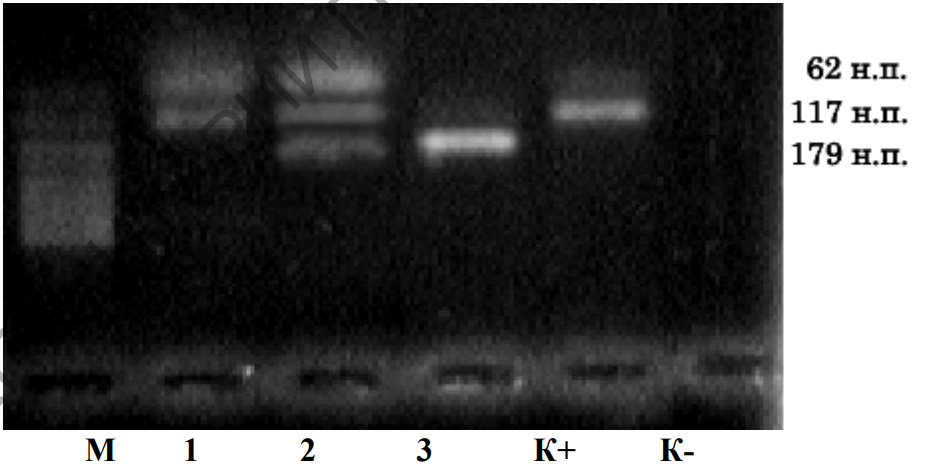
Рисунок 10 – Электрофореграмма продуктов рестрикции по гену TYR
Из рисунка 10 видно, что образец 1 включает 2 фракции (62 н.п. и 117 н.п.), что возможно только для особей с генотипом СС. Образец 2 содержит 3 фракции (62 н.п., 117 н.п. и 179 н.п.), что говорит о гетерозиготном генотипе Ccs. Что касается 3 образца, то там присутствует только одна фракция (179 н.п.), что характеризует гомозиготу cscs.
В результате проведенных исследований были отработаны и апробированы методы молекулярно-генетической идентификации особей F. catus акромеланистических окрасов. Позволяющие четко выявлять гетерозиготных особей (генотип Ccs), которые фенотипически неотличимы от особей с генотипом СС.
Работа проводилась в рамках темы ГБ 21-34 «Фаунистические и физиологические исследования животных и человека Гомельского региона».
Литература
- Гончаренко, Г. Г. Генетика. Анализ наследственных закономерностей на генах меха кошек
Felis catus : практическое пособие / Г. Г. Гончаренко, С. А. Зятьков. – Гомель : ГГУ им. Ф. Скорины, - – 108 с.
- О’Брайен, С. Генетика кошки / С. О’Брайен, Р. Робинсон, А. С. Графодатский, А. В. Таранин. –
Новосибирск : Наука, 1993. – 212 с. - Searle, A. G. Comparative genetics of coat color in mammals / A. G. Searle. – London : Logos P.;
New York : Academic P., 1968. – 308 p. - V. del Marmol, V. Tyrosinase and related proteins in mammalian pigmentation / V. del Marmol,
F. Beermann // FEBS Lett. – 1996. – Vol. 381. – P. 165–168. - Hearing, V. J. Enzymatic control of pigmentation in mammals / V. J. Hearing, K. Tsukamoto //
FASEB J. – 1991. – Vol. 5. – P. 2902–2909. - Pawelek, J. M. The enzymology of melanogenesis / J. M. Pawelek, A. K. Chakraborty. //
Pigmentary System. – New York : Oxford University Press, 1998. – P. 391–400. - Зятьков, С. А. Характер наследования и механизмы взаимодействия генов окраски меха у
домашних кошек / С. А. Зятьков // Известия ГГУ им. Ф. Скорины. – 2007. – № 6 (45). – С. 81–86. - Зятьков, С. А. Молекулярно-генетическая дактилоскопия особей Felis catus акромеланистических окрасов / С. А. Зятьков, И. В. Курако // Ростовский научный журнал. – 2017. – № 12 (50). –
С. 362–366. - Lyons, L. A. Tyrosinase mutations associated with Siamese and Burmese patterns in the domestic
cat (Felis catus) / L. A. Lyons, D. L. Imes, H. C. Rah, R. A. Grahn // Anim. Genet. – 2005. – № 36 (2). –
P. 119–126. - Schmidt-Kuntzel, A. Tyrosinase and Tyrosinase Related Protein 1 Alleles Specify Domestic Cat
Coat Color Phenotypes of the albino and brown Loci / A. Schmidt-Kuntzel, E. Eizirik, S. J. O’Brien,
M. Menotti-Raymond // Journal of Heredity. – 2005. – Vol. 96 (4). – P. 289–301. - Imes, D. L. Albinism in the domestic cat (Felis catus) is associated with a tyrosinase (TYR)
mutation / D. L. Imes, L. A. Geary, R. A. Grahn, L. A. Lyons // Animal Genetics. – 2006. – Vol. 37. –
P. 175–178. - Yu, Y. Mocha tyrosinase variant : a new flavour of cat coat coloration / Y. Yu, R. A. Grahn,
L. A. Lyons // Animal Genetics. – 2019. – Vol. 50. – P. 182–186. - O’Brien, S. J. The Feline Genome Project / S. J. O’Brien, M. Menotti-Raymond, W. J. Murphy,
N. Yuhki // Annu. Rev. Genet. – 2002. – Vol. 36. – P. 657–686. - Pontius, J. U. Initial sequence and comparative analysis of the cat genome / J. U. Pontius [et al.] //
Genome Res. – 2007. – Vol. 17. – P. 1675–1689. - Cho, K. W. A proposed nomenclature of the domestic cat karyotype / K. W. Cho [et al.] //
Cytogenet Cell Genet. – 1997. – Vol. 79. – P. 71–78.


