
Energy and protein needs of cats for maintenance, gestation and lactation
B Wichert1*, L Schade1, S Gebert2, B Bucher2, B Zottmaier1, C Wenk2, M Wanner1
Journal of Feline Medicine and Surgery (2009) 11, 808e815
1 Institute of Animal Nutrition,
Vetsuisse Faculty University Zurich, Switzerland 2 Institute of Animal Sciences, Swiss
Federal Institute of Technology
Zurich, Switzerland
Date accepted: 12 February 2009
In the present investigation, data on the energy intakes and energy needs, as well as protein and fat accretion, of queens during pregnancy, during lactation and after lactation are given. Eleven adult cats were used as experimental animals. Data were collected during the fourth and seventh week of pregnancy, the second and sixth week of lactation and the second and sixth week after lactation. The cats were fed dry kitten food. During gestation and after lactation, all measurements were performed with respiration chambers. During lactation, balance trials without respiration chambers were performed. Body weight was measured and nitrogen, carbon and energy balances were calculated. From these, protein and fat accretion, as well as the metabolisable energy intake, was calculated. The weight gain during gestation was linearly independent of the number of kittens. During lactation, all cats lost weight; nevertheless, all cats except one were heavier 2 weeks after lactation than at mating. The energy intake of the cats during gestation was 1.8 times the maintenance requirement in the fourth week and two times maintenance requirement in the seventh week, and these energy intakes differed greatly among individuals. The energy intake of the cats during lactation was clearly higher than that recommended by National Research Council (NRC)1, whereas the recommended protein intake in the second week of lactation was met. As the calculated protein balance was negative, the NRC recommendation for protein intake seems to be too low. In comparison to previous data, the cats showed a higher energy intake during lactation (median 502 kJ/kg BW/d, second week lactation), and the weight loss was much lower. Further investigations on pregnant and lactating cats are necessary to complete the database.
Сurrently, there is little data available on the energy requirements of queens during gestation and lactation. During gestation, cats increase their body weight (BW) because of the growth of the fo et uses and to build up body reserves.2 Queens gain weight linearly during pregnancy beginning at the first day of pregnancy.2 This finding is contrary to bitches that show the main increase in BW after the 28th day of gestation.3 The weight gain of the queens includes fo et al, placental and associated tissues, as well as body reserves of the queen,1 as they need reserves for lactation. Loveridge and Rivers4 observed an increase in BW in pregnant cats of about 32%. Twenty-four hours post partum, the queens weighed between 19% and 26% more than before mating. At the end of lactation, the BW of all of the cats was in a range of 2% of the pre-mating weight. Queens with bigger litters lost weight faster.4 Loveridge2 and Zottmann5 also observed that during the seventh to ninth weeks of lactation, the queens lost weight regardless of their diet. The weight loss in the first 4 weeks of lactation was also confirmed by Hendriks and Wamberg.6 These authors did not give data concerning weeks five to eight of lactation. Loveridge2 described a BW change for the whole period of pregnancy and lactation that was between an increase of 18 g (0.6%) in BW for a queen with one kitten and a decrease of 29 g (0.8%) in BW for a queen with five kittens. The BW changes of all other queens were in between these values.
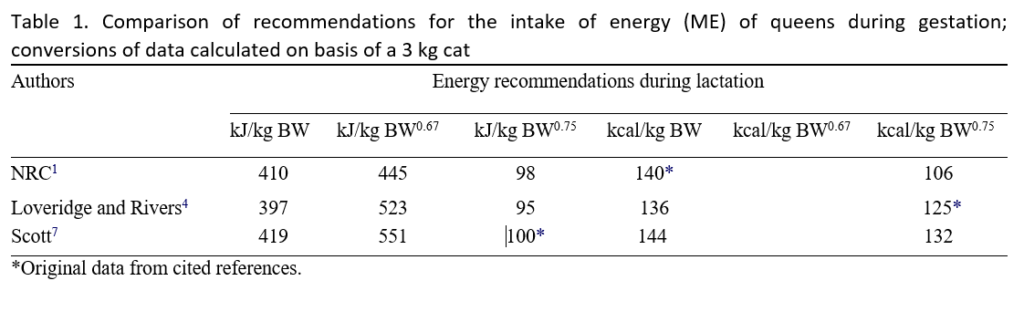
Scott7 postulated an energy requirement of 100 kcal ME/kg BW for pregnant cats. For the calculation of metabolisable energy (ME), modified Atwater factors were used. Loveridge and Rivers4 gave a similar requirement of 90 kcal ME/kg BW. Based on the data of the cited authors, the NRC1 recommends an energy requirement of pregnant cats of about 140 kcal ME/ kg BW0.67. Table 1 shows a comparison of the different data on energy requirements of pregnant queens. However, as the ME content of pet food is often overestimated,8 the NRC1 recommends a safety margin, which means a slightly higher allowance than 140 kcal ME/ kg BW0.67 should be used when feeding pregnant cats.
According to Loveridge and Rivers,4 the energy intake in the first week of lactation differed between 58 kcal ME/kg BW/d with one kitten and 118 kcal ME/kg BW/d with five kittens. It increased until the sixth week of lactation to 97 kcal ME/kg BW/d with one kitten and 354 kcal ME/kg BW/d with five kittens. In their study, the kittens began to feed on the same food as their mothers starting in the third or fourth week of life. Therefore, their energy intake is included in the given data.
According to Scott,7 a lactating queen needs 1046 kJ ME/kg BW/d (¼250 kcal ME/kg BW/d). For a cat raising three kittens in the third week of lactation, the NRC1 recommends an energy intake (ME) of 100 kcal/kg BW0.67 (for maintenance) þ 60 BW L (where L is a factor for the week of lactation, in the third week L¼ 1.2). This means that for a cat of 3 kg BW, the energy requirement is 425 kcal ME/d (1778 kJ ME/d). The energy requirement of lactating cats at the peak of lactation in the third lactation week (810 kJ ME/kg BW/d ¼ 193 kcal ME/kg BW/d; for a 3 kg cat: 579 k cal ME/d or 2430 kJ ME/d) was factorially calculated byKienzle.9 Thismeansthattheenergyformaintenance requirements, as well as the energy output with milk (energy content of milk multiplied with milk yield), for different weeks of lactation was calculated, and the utilisation of ME was estimated from knowledge of other mammals like the dog,9 as there is no data on the utilisation of ME for milk production in cats.10
In earlier experiments, the queens did not reach the calculated requirements for lactation, and as a result, they lost weight.2,5,6 The aim of the present study was to measure the energy need sof queens, as wel last heprotein and fatac cretion, during different stages, from mating to 6 weeks after
lactation. Therefore, the respiration technique was used whenever possible (not during lactation).
Materials and methods
In the present investigation, 11 adult cats with a mean BW of 3.0 0.3 kg and an average age of 1.3 0.3 years were measured before mating (adult). The cats were reared in the laboratory for research purposes by the institute. They were maintained according to the animal welfare legislation of Switzerland. During the fourth week (first half of gestation) and seventh week of gestation (second half of gestation), 9 and 10 cats, respectively, were
measured. In the second week (peak of lactation) and sixth week of lactation (decreasing milk yield), six and five cats, respectively, were measured. In the second week (shortly after weaning) and sixth week after weaning (cats should have come back to pre-mating status), six cats each were measured. The number of cats differed between the measurements because some cats had diarrhoea during some of the measurements, and, therefore, that data could not be evaluated. Furthermore, one cat lost the kittens on the first day of lactation, and in one queen, the kittens had to be weaned earlier. For all cats, this was the first pregnancy and lactation. The measurements during gestation and after weaning were performed for 5 days each in respiration chambers (1.4 m 1.0 m 0.9 m) shown in Fig 1, for 22.5 h per day. The chambers were open circuit units. Inside the chambers the air was continuously renewed by external air as well as recycled air of the chambers. The CO2 and O2 concentrations of the chambers air did not differ more than 1% from the external air. The temperature of recycled air passed air condition before entering the chambers. The chambers were under slight vacuum to prevent loss of air through small leaks. The flow rate was 3.5 m3/h.
The first day was used for adaptation to the new environment. Only the measurements from the second to fifth days were used. In the second and sixth week of lactation, only the food intake and complete collection of faeces and urine of the queens were measured for 7 days. The queens and their litters were kept together during this time. The kittens received additional feeding from the third week. In the sixth week of lactation, the queens and kittens were fed separately and used separate litter boxes. The food and litter boxes, were placed, that either only the queens or the kittens could reach them.
Fig 1. The respiration chambers.

During the time in the respiration chambers, the O2 consumption and the CO2 production of the cats were determined with help of the respiration chambers of the ETH Zu¨rich.11 Measurements of O2 and CO2 (and CH4) contents of the air leaving the chamber as well as the temperature were monitored and recorded through 100 s intervals for each chamber. The concentration of O2 was measured by a paragmetic analyser (Oxymat 3, Siemens
AG, Dietikon, Switzerland) in a range of 19e21%. Concentration of CO2 and CH4 was analysed by an infrared method (Ultramat 2, Siemens AG, Dietikon, Switzerland). All data were collected automatically with help of an EPROM (Leybold-Hera¨us AG, Zu¨rich, Switzerland). The signals of each analyser were transferred through an A/D converter and directly recorded by PC. All data were stored as an arithmetic mean of each measuring interval. Before each period of measurements in the respiration chambers the recovery rate for CO2 and O2 was determined. Therefore, propane gas was burned inside the chambers. The simultaneous vapourisation of water simulated an animal experiment. Assuming total burning of the gases, CO2 production as well as O2 consumption was determined from the difference in weight of the propane gas. The recovery rate was at least 90%. Every day between 9.00 and 10.30 am, the chambers were opened. During this time, the chambers were cleaned, samples were taken and the cats were given time out of the chambers and the instruments for the measurements of the gases were calibrated with standard calibration gas (1016 ppm CH4, 1.02% CO2, 19.52% O2) and N2 (quality 5.0; Pan Gas, Dagmarsellen, Switzerland). The cats were exposed to light for 16 h per day between 6.00 and 22.00, similar to household cats in the summertime.
During thewhole experiment, the catswerefed commercial dry food for kittens (Biomill kitten, Biomill AG, Granges Marnand, Switzerland) with the addition of 2% celite (Karl Roth GmbH, Karlsruhe, Deutschland) as an internal marker (Table 2). The food intake was measured daily. Faeces and urine were collected total and separated with help of special litter boxes that allowed the cooling of the urine.12 The litter boxes contained a non-absorbing polypropylene cat litter. Faeces and urine were lyophilised before analysis. The crude nutrients were analysed in the food and faeces. Food (dried), urine and faeces (lyophilised) were used for gross energy determination by bomb calorimetry (IKA calorimeter C2000 basic; IKA-Werke GmbH, Staufen, Germany). The dried food, lyophilised faeces and urine (defrosted only) were analysed for carbon (C) and nitrogen (N) with a CN-analyser (Leco CN-2000; Leco Instrumente GmbH, Mo¨nchengladbach, Germany). N, C and energy balances were calculated for the following measuring periods: adult, fourth and seventh week of gestation and second and sixth week after weaning. Digestibility of crude nutrients and energy was determined: digestibility (%) of crude nutrients (cn) ¼ (cn food cn faeces)/cn food 100. The energy balances were calculated from Brouwer13:

In lactating cats energy intake (ME) was calculated
(intake gross energyfood gross energyfaeces þ gross energyurine ¼ intake ME).
The energy requirement was calculated from the NRC guidelines.1 The data also account for the number of kittens. The protein balance in lactating cats was calculated from the retained protein minus the protein content of milk (8.1% crude protein (CP)) multiplied by the calculated milk yield depending on the litter size. The milk yield in the second week of lactation was 1.41% of the queen BW with 2 kittens, 5.36% of the queen BW with three to four kittens and 8.04% of the queen BW with six kittens. The milk yield in the sixth week of lactation was 1.27% of the queen BW with 2 kittens, 4.71% of the queen BW with three to four kittens and 5.69% of the queen BW with six kittens.5
Normally distributed data are given as mean values and standard deviation (SD), or if outliers were present the median with minimum and maximum, was calculated. A repeated analysis of variance (ANOVA) was performed with the group as a cofactor. The energy requirements of the cats during different stages of life were calculated by linear regression analyses from the intake of ME and energy balance.
Results
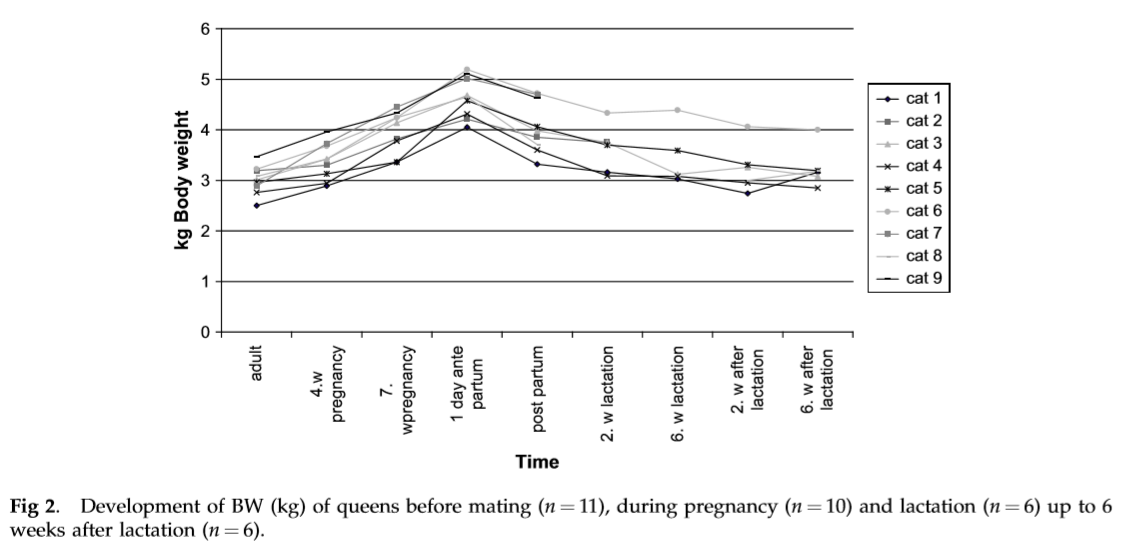
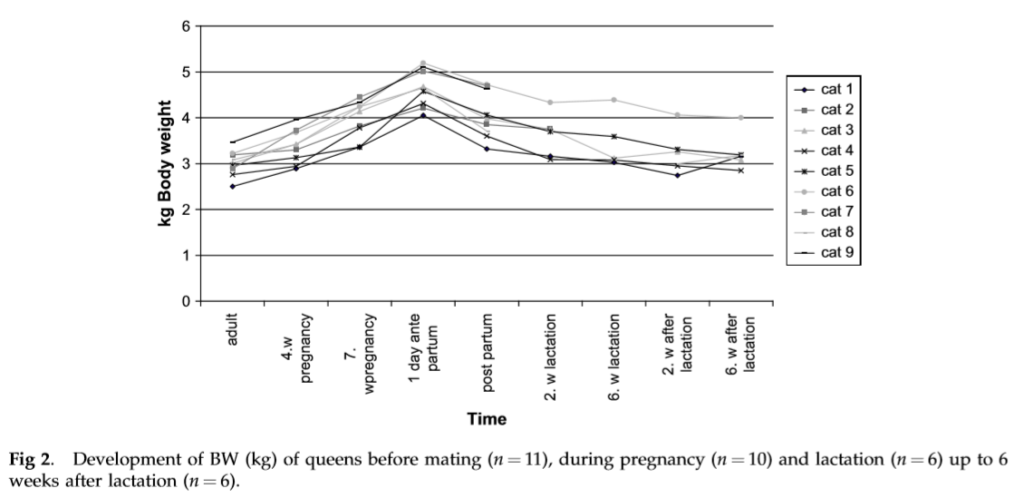
The 11 cats considered ‘adults’ weighed on average 3.0 0.3 kg. During pregnancy, the nine measured cats showed a median increase in BW of 14.1% (min 3.5% and max 29.1%) up to the fourth week of gestation and 34.4% (min 13.5% and max 54%) up to the seventh week of gestation. The weight gain during gestation of all cats showed a nearly linear increase (Fig 1). The cats gave birth to between two and six kittens (2 kittens (k) ¼ 1 cat (c), 3 k ¼ 1 c, 4 k ¼ 2 c, 5 k ¼ 4 c, and 6 k ¼ 1 c). The median weight gain of the queens was 1.6 kg (min 1.0 kgandmax2.1 kg).The cat with thesmallest weight gain (1.0 kg) had four kittens, and the cat with the largest weight gain (2.1 kg) had three kittens.
Table 2. Proximateanalysis,GE,andHCl-insoluble ash of the diet, values in % original matter, versus kJ (GE)

Table 3. Apparent digestibility of crude nutrients (%) during adult measurement (n¼ 11), fourth week of gestation (n¼ 8) and seventh week of gestation (n¼ 9)
At parturition, the cats lost a median 520 g (min 320 g and max 940 g) of BW, which corresponds to a weight loss of 12.8% (min 6.8% and max 25.3%). During lactation, all cats lost weight in comparison to their weight after parturition. Nevertheless, two cats gained weight again during the measuring periods in the second week of lactation (0.07 and 0.28 kg), and in the sixth week of lactation, three cats gained weight (0.07e0.1 kg) again. Two weeks after lactation, 5/6 cats were heavier than before mating (min 2.6% of BW adult and max þ26.1% of BW adult). In the sixth week after lactation, all the cats were between 2.3% and 26.4% heavier than before mating. The kittens showed an average dailyweightgainof12.5 2.3 gduringthefirst9weeks of age. The weight gain of the kittens was not dependent on litter size.
The median energy intake (ME) during the fourth and seventh week of pregnancy was 389 kJ/kg BW/ d (min 276 and max 433 kJ/kg BW/d) and 361 kJ/ kg BW/d (min 308 and max 453 kJ/kg BW/d), respectively. In com paris onto their main te nance requirement, the energy intake per cat in the fourth week of pregnancy was 1.8 times higher, and in the seventh week of pregnancy, it was twice the maintenance energy requirement. The metabolisability of energy was
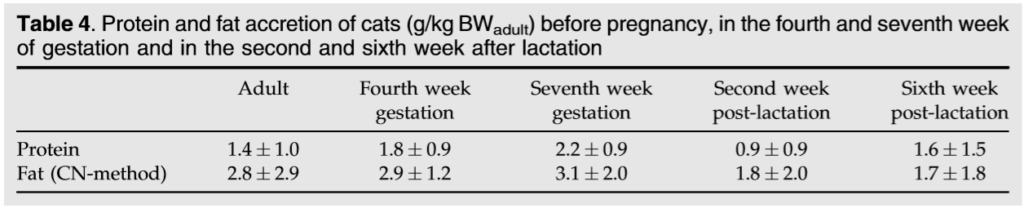
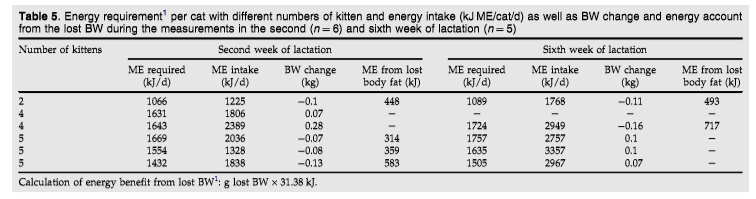
0.85 0.02 kJ/kJ in the fourth week of gestation and 0.84 0.03 kJ/kJ in the seventh week of gestation. The digestibility of CP, crude fat (SF), crude fibre (CF), and nitrogen free extracts (NfE), as well as gross energy (GE), were similar during the adult measurements, the fourth and seventh week of pregnancy and the second and sixth week of lactation (Table 3). The cats retained a median of 126 kJ/kg BW/d (min 63 and max 222 kJ/kg BW/d) energy in the fourth week of gestation, and inth eseventh week of gestation, a comparable amount at 144 kJ/kg BW/d (min 75 and max 268 kJ/ kg BW/d) was retained. No significant differences in protein accretion and fat accretion were observed between non-pregnant cats, which were slightly variable in weight during the measurement in the respiration chambers, and cats during gestation (Table 4). In the second week of lactation, the lactating cats showed a highly variable energy intake. The median energy intake was 502 ME/kg BW/d in the second week of
lactation (min 335 kJ ME/kg BW/d with two kittens and max 599 kJ ME/kg BW/d with five kittens) (Table 6). The cats that had at least three kittens showed a median energy intake of 520 kJ ME/kg BW/d during this time (Table 5). In the sixth week of lactation, the queens had a median energy intake of 746 kJ ME/ kg BW/d (min 422 kJ ME/kg BW/d with four kittens and max 849.2 kJ ME/kg BW/d with five kittens). In the second week of lactation, all the cats except one showed a higher energy intake (Table 6) than the calculated NRC recommendations.1 In the sixth week of lactation (Table 6), all the cats consumed significantly more energy than recommended by the NRC.1
The protein intake of the cats in the second week of lactation reached a median 17.0 g/kg BW0.67 (min 11.3 g/kg BW0.67 and max 20.9 g/kg BW0.67), whereas in the sixth week of lactation, the median intake of protein was 21.2 g/kg BW0.67 (min 13.6 g/kg BW0.67 and max 28.2 g/kg BW0.67). The nitrogen retained before the subtraction of milk was 0.5 0.2 g/kg BW adult/d in the second week of lactation and
0.8 0.4 g/kg BWadult/d in the sixth week of lactation. In the second week of lactation, the retained protein after the subtraction of milk protein (milk yield calculated) was 3.4 g protein/d for a cat with two kittens; for cats with four kittens, the retained protein lost was a minimum of 1.55 g protein/d and a maximum of
9.9 g protein/d. The cats with five kittens lost a minimum of 3.6 g protein/d and a maximum of 8 g protein/d. In the sixth week of lactation, the retained protein in all cats averaged between 2.5 g protein/d (four kittens) and 16.6 g protein/d (five kittens).
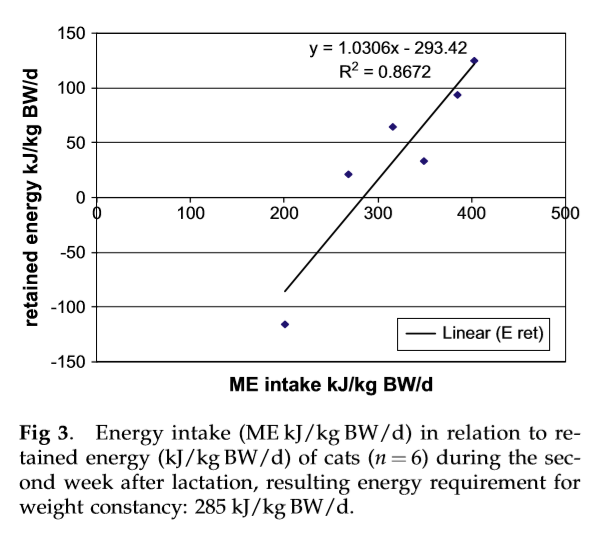
During the measurement in the second week after lactation, five cats retained energy, and only one cat showed a negative energy balance (112.3 kJ/kg BW adult/d). The energy requirement of the cats for weight constancy, as determined by regression analyses (r2 ¼ 0.89), in the second week after lactation was 285 kJ/kg BW/d (Figs 2, 3). In the sixth week after lactation, the energy requirement of the cats for weight constancy was similar at 261 kJ/kg BW/d (r2 ¼ 0.97).
Discussion
During gestation, the cats were fed ad libitum. Their weight gain during gestation time was nearly linear. This is in agreement with Loveridge,2 who observed a steady increase in BW beginning with the first day of pregnancy. The total weight gain of the queens during gestation varied between individuals, but the average weight gain was similar to that measured by LoveridgeandRivers.4 The parturition cause da weight loss of 12.3 5.2% of the BW of the cats in the present investigation, which is also in good agreement with Loveridge and Rivers,4 where the cats lost between 6% and 13% of the BWat the day of parturition. In contrast to the studies of Loveridge and Rivers4 and Zottmann,5 most of the queens (5/6) of the present study were heavier 2 weeks after lactation than before mating. At 2 and 6 weeks after lactation, only one cat that nursed six kittens showed a BW in the range of 2.6% and þ2.3% compared to her BW at mating, which is in agreement with the data of Loveridge and Rivers.4 In the studies of Loveridge2 and Loveridge and Rivers,4 the feeding regimen is not given in detail, and the cats of Zottmann5 were fed with a combination of dry food and canned food. It can be speculated that the adlibitum feeding of dry food, which was relatively high in energy content, led to a higher intake of food and, as a consequence, energy, especially during lactation. Another possibility would be that the cats of the presentc study that show eda similar weight gain during pregnancy and a similar weight loss through parturition had a lower resting metabolic rate in comparison to the cats of the study of Zottmann,5 or perhaps they were less active and maybe less stressed during the lactation time. A situation of less stress, at least in comparison to the study of Zottmann,5 could be explained by the milking of the queens and, for this reason, the temporary separation of the kittens in the study of Zottmann.5 Therefore, the queens of the present study would have needed less energy, and consequently, the weight loss was lower than in the other studies.2,4,5 The energy intake of the queens in the fourth and seventh week of gestation was in the same range as that observed by Loveridge and Rivers,4 the NRC1 and Scott7 (Table 1).
The metabolis ability of energy during gestation, 0.85 0.02 kJ/kJ and 0.84 0.03 kJ/kJ, corresponded to a value of 0.85 0.03 kJ/kJ measured in the cats during the adult measurement.12 It cannot be elucidated from the present data whether the tendency to a lower metabolis ability of energy in the seventh week of pregnancy was caused by the tendency to a lower NfE digestibility or if simply the differences were too low to be seen in the respiration measurements. The retained energy per kg BW/d did not differ between the fourth and seventh week of pregnancy, which fits the estimation that cats gain weight linearly during gestation. The gained weight consisted of a higher amount of fat than of protein. This would confirm the speculation of the NRC1 that the cats build up body reserves as a reserve for lactation. In addition, in sows, a so-called pregnancy ecommended by the NRC.1 The recommendations include the expectance that all queens lose BW during lactation. It was thought that at least the queens with larger litter sizes would not be able to reach their energy requirements. It has been speculated that they use the body stores builtup during pregnancy to provide energy for milk production.
Nevertheless,2/6 cats increased BW during early lactation(Table5).This could be ex plain edit cats wit hone or two kittens were able to take up as much energy as needed for milk production. However, in the present investigation, this was not true as the cat with only two kittens lost BW, but the ones with four kittens did not. In the second week of lactation, only one cat showed a lower ME intake than recommended by the NRC.1 This cat was not the one with the maximum decrease in BW.
The very individual results of energy intake and weight loss of the queens in comparison to the recommended energy intake indicate that other influences, such as activity and stress of the cats, are import ant factors for their real energy needs. As the milk yield of the queens was not measured, differences in milk yield could also cause the differences in weight loss, independent of energy intake. However, the weight development of the kittens was very similar between the kittens of the present study, as well as to the kittens of the study of Zottmann.5 Therefore, it can be assumed that the milk yield of the mothers per kitten was also similar inside the present study. In comparison to the study of Zottmann,5 the kittens in the present study began to eat additional food earlier. Consequently, it cannot be excluded that the total milk yieldof the queen sof the present study was lower.
The calculated protein balance also showed individually different results in the cats and no relation to litter size, and therewith, no differences in milk yield or energy and protein intake were observed. This means that the changes in body composition differed between the cats. Although all cats except the one with only two kittens consumed the required amount of CP,1 all the cats showed a negative protein balance in the second week of lactation. It follows that the protein recommendation of the NRC1 is too low for lactating cats. From this, it can be assumed that the food for lactating cats should be high in protein content. From the present investigation, it cannot be determined if it is possible to get a sufficient supply of protein for lactating cats using a food higher than 33% of protein, as the one used here. In the sixth week of lactation, which is after the peak of lactation, most of the cats showed positive protein balances again, if the calculated milk yield is in agreement with the one of the queens of the present investigation. As the kittens were fed additional food beginning in the third week of lactation, the milk yield of the queens could have been lower than in the investigation of Zottmann,5 where the additional feeding was started at the 25th day of life.
Up to the sixth week after lactation, two cats normalised their BW, but the other cats stayed heavier than before mating. For this reason, it is difficult to determine their energy requirements with the help of regression analyses. To keep their actual BW constant, the cats needed 285 kJ/kg BW/d in the second week after lactation and similarly 261 kJ/kg BW/d in the sixth week after lactation, which was a little higher than during the adult measurement that was done before mating (238 kJ/kg BW/d16). To the author’s knowledge, nothing is known about body composition of the cats. It is assumed that normally during lactation fat and fat free mass are used for milk production.1 This would lead to changes in body composition. But in the present investigation, most of the cats were heavier after lactation than before mating. Therefore, it is likely that they gained fat tissue during gestation and maybe also during lactation. Normally, the energy needs of overweight cats are lower per kg BW.17 This was not true for the cats during the second and sixth week after lactation in the present investigation. As the cats were on average 1.3 years at the beginning of the studyie, they were full grown,18 the higher energy need could be caused by the higher activity of the cats after weaning.
In conclusion, the knowledge about the energy and protein needs of cats during lactation is very scarce. Further investigations with different kinds of diet or feeding regimes need to be performed to complete our knowledge of energy and protein metabolism, as well as energy and protein requirements, in cats.
Acknowledgement
The authors thank Biomill SA for the support of the study
References
- National Research Council. Nutrient requirements ofdogs and cats. Washington, DC: National Academic Press, 2006, 28e48, 114e20, 364e70.
- Loveridge GG. Bodyweight changes and energy intakeof cats during gestation and lactation. Anim Technol 1986; 37: 7e15.
- Meyer H, Dammers C,Kienzle E. Ko¨rperzusammensetzung neugeborener Welpen und Na¨hrstoffbedarf tragender Hu¨ndinnen. Adv Anim Physiol Anim Nutr 1985; 16: 7e25.
- Loveridge GG, Rivers JPW. Bodyweight changes and energy intakes of cats during pregnancy and lactation. In: Burger IH, Rivers JPW, eds. Nutrition of the dog and cat, Waltham symposium 7. Cambridge: University Press, 1989: 113e32.
- Zottmann B. Untersuchungen zur Milchleistung undMilchzusammensetzung der Katze (Felis catus). Diss Med Vet, Mu¨nchen, 1997.
- Hendriks WH, Wamberg S. Milk intake of suckling kittens remains relatively constant from one to four weeks of age. J Nutr 2000; 77: 16e23.
- Scott PP. The cat: nutrition. UFAW handbook on the careand management of laboratory animals, 3rd edn. Edinburgh: Livingstone Ltd, 1966: 505e67.
- Kendall PT, Burger IH, Smith PM. Methods of estimationof the metabolisable energy content of cat foods. Feline Pract 1985; 15: 38e44.
- Kienzle E. Factorial calculation of nutrient requirementsin lactating queens. J Nutr 1998; 128: 2609Se14.
- Scantlebury M, Butterwick R, Speakmann JR. Energeticsof lactation in domestic dog (Canis familiaris) breeds of two sizes. Comp Biochem Physiol Part A197e210.
- Hadorn R. Einfluss unterschiedlicher nahrungsfasertra¨ger (soja- und hirseschalen) im vergleich zu weizenquellsta¨rke auf die Na¨hrstoff- und energiebewertung von wachsenden schweinen und broilern. Diss, ETH Zu¨rich, 1994.
- Schade L. Untersuchungen zum energiestoffwechsel vontra¨chtigen katzen. Diss Med Vet, Zu¨rich, 2006.
- Brouwer E. Report of subcommittee on constants and factors. In: Blaxter KL, ed. Proceedings of the 3rd Symposium on Energy Metabolism. Vol. 11, Troon, Scotland: EAAP Publ., 1965: 441e3.
- Noblet J, Dourmad JY, Etienne M. Energy utilization inpregnant and lactating sows: modelling of energy requirements. J Anim Sci 1990; 68: 562e72.
- Shurson GC, Libal GW, Crenshaw J, et al. Impact ofenergy intake and pregnancy status on rate and efficiency of gain and backfat changes of sows postweaning. J Anim Sci 2003; 81: 209e16.
- Wichert B, Mu¨ller L, Gebert S, Wenk C, Wanner M. Additional data on energy requirements of young adult cats measured by indirect calorimetry. J Anim Physiol Anim Nutr 2007; 91: 278e81.
- Edstadtler-Pietsch G. Untersuchungen zum Energiebedarf von Katzen. Diss Med Vet, Mu¨nchen, 2003.
- Root MV, Johnston SD, Olson PN. Effect of pre puberal and post puberal gonadectomy on heat production measured by indirect calorimetry in male and female domestic cats. Am J Vet Res 1996; 57: 371e4.



